What Does the Multiregional Continuity Thesis Propose
Early Modern Homo sapiens
All people today are classified as Homo sapiens . Our species of humans first began to evolve nearly 200,000 years ago in association with technologies not unlike those of the early Neandertals. It is now clear that early Homo sapiens, or modern humans, did not come after the Neandertals but were their contemporaries. However, it is likely that both modern humans and Neandertals descended from Homo heidelbergensis.
Compared to the Neandertals and other late archaic humans, modern humans generally have more delicate skeletons. Their skulls are more rounded and their brow ridges generally protrude much less. They rarely have the occipital buns found on the back of Neandertal skulls. They also have relatively high foreheads, smaller faces, and pointed chins.
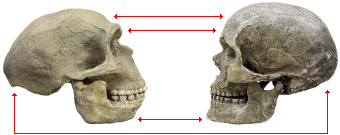 | |
| Neandertal | modern Homo sapiens |
The first fossils of early modern humans to be identified were found in 1868 at the 27,000-23,000 year old Cro-Magnon rock shelter site near the village of Les Eyzies ![]() in southwestern France. They were subsequently named the Cro-Magnon
in southwestern France. They were subsequently named the Cro-Magnon ![]() people. They were very similar in appearance to modern Europeans. Males were 5 feet 4 inches to 6 feet tall (1.6-1.8 m.) That was 4-12 inches (10-31 cm.) taller than Neandertals. Their skeletons and musculature generally were less massive than the Neandertals. The Cro-Magnon had broad, small faces with pointed chins and high foreheads. Their cranial capacities were up to 1590 cm 3 , which is relatively large even for people today.
people. They were very similar in appearance to modern Europeans. Males were 5 feet 4 inches to 6 feet tall (1.6-1.8 m.) That was 4-12 inches (10-31 cm.) taller than Neandertals. Their skeletons and musculature generally were less massive than the Neandertals. The Cro-Magnon had broad, small faces with pointed chins and high foreheads. Their cranial capacities were up to 1590 cm 3 , which is relatively large even for people today.
Origins of Modern Humans
Current data suggest that modern humans evolved from archaic humans primarily in East Africa. A 195,000 year old fossil from the Omo 1 site in Ethiopia shows the beginnings of the skull changes that we associate with modern people, including a rounded skull case and possibly a projecting chin. A 160,000 year old skull from the Herto site in the Middle Awash area of Ethiopia also seems to be at the early stages of this transition. It had the rounded skull case but retained the large brow ridges of archaic humans. Somewhat more advanced transitional forms have been found at Laetoli in Tanzania dating to about 120,000 years ago. By 115,000 years ago, early modern humans had expanded their range to South Africa and into Southwest Asia (Israel) shortly after 100,000 years ago. There is no reliable evidence of modern humans elsewhere in the Old World until 60,000-40,000 years ago, during a short temperate period in the midst of the last ice age.
| Important Early Modern Homo sapiens Sites | |||||||||||||||||||||||||||||||||||||||||||||||||
| |
| ||||||||||||||||||||||||||||||||||||||||||||||||
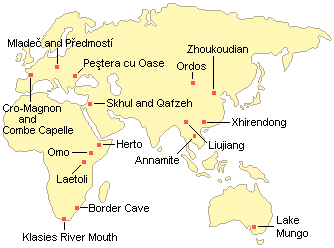
| Note: Artifactual evidence indicates that modern humans were in Europe by at least 40,000 and possibly as early as 46,000 years ago. Dating of the earliest modern human fossils in Asia is less secure, but it is likely that they were present there by at least 60,000 years ago and possibly 100,000 years ago. |
It would seem from these dates that the location of initial modern Homo sapiens evolution and the direction of their dispersion from that area is obvious. That is not the case. Since the early 1980's, there have been two leading contradictory models that attempt to explain modern human evolution--the replacement model and the regional continuity model.
The replacement model of Christopher Stringer and Peter Andrews proposes that modern humans evolved from archaic humans 200,000-150,000 years ago only in Africa and then some of them migrated into the rest of the Old World replacing all of the Neandertals and other late archaic humans beginning around 60,000-40,000 years ago or somewhat earlier. If this interpretation of the fossil record is correct, all people today share a relatively modern African ancestry. All other lines of humans that had descended from Homo erectus presumably became extinct. From this view, the regional anatomical differences that we now see among humans are recent developments--evolving mostly in the last 40,000 years. This hypothesis is also referred to as the "out of Africa", "Noah's ark", and "African replacement" model.
The regional continuity model (or multiregional evolution model) advocated by Milford Wolpoff proposes that modern humans evolved more or less simultaneously in all major regions of the Old World from local archaic humans. For example, modern Chinese are seen as having evolved from Chinese archaic humans and ultimately from Chinese Homo erectus. This would mean that the Chinese and some other peoples in the Old World have great antiquity in place. Supporters of this model believe that the ultimate common ancestor of all modern people was an early Homo erectus in Africa who lived at least 1.8 million years ago. It is further suggested that since then there was sufficient gene flow between Europe, Africa, and Asia to prevent long-term reproductive isolation and the subsequent evolution of distinct regional species. It is argued that intermittent contact between people of these distant areas would have kept the human line a single species at any one time. However, regional varieties, or subspecies, of humans are expected to have existed.

Replacement Model Arguments
There are two sources of evidence supporting the replacement model--the fossil record and DNA. So far, the earliest finds of modern Homo sapiens skeletons come from Africa. They date to nearly 200,000 years ago on that continent. They appear in Southwest Asia around 100,000 years ago and elsewhere in the Old World by 60,000-40,000 years ago. Unless modern human remains dating to 200,000 years ago or earlier are found in Europe or East Asia, it would seem that the replacement model better explains the fossil data for those regions. However, the DNA data supporting a replacement are more problematical.
Beginning in the 1980's, Rebecca Cann, at the University of California, argued that the geographic region in which modern people have lived the longest should have the greatest amount of genetic diversity today. Through comparisons of mitochondrial DNA sequences from living people throughout the world, she concluded that Africa has the greatest genetic diversity and, therefore, must be the homeland of all modern humans. Assuming a specific, constant rate of mutation, she further concluded that the common ancestor of modern people was a woman living about 200,000 years ago in Africa. This supposed predecessor was dubbed "mitochondrial Eve" ![]() . More recent genetic research at the University of Chicago and Yale University lends support to the replacement model. It has shown that variations in the DNA of the Y chromosome and chromosome 12 also have the greatest diversity among Africans today. John Relethford and other critics of the replacement model have pointed out that Africa could have had the greatest diversity in DNA simply because there were more people living there during the last several hundred thousand years. This would leave open the possibility that Africa was not necessarily the only homeland of modern humans.
. More recent genetic research at the University of Chicago and Yale University lends support to the replacement model. It has shown that variations in the DNA of the Y chromosome and chromosome 12 also have the greatest diversity among Africans today. John Relethford and other critics of the replacement model have pointed out that Africa could have had the greatest diversity in DNA simply because there were more people living there during the last several hundred thousand years. This would leave open the possibility that Africa was not necessarily the only homeland of modern humans.
Critics of the genetic argument for the replacement model also point out that the rate of mutation used for the "molecular clock" is not necessarily constant, which makes the 200,000 year date for "mitochondrial Eve" unreliable. The rate of inheritable mutations for a species or a population can vary due to a number of factors including generation time, the efficiency of DNA repair within cells, ambient temperature, and varying amounts of natural environmental mutagens. In addition, some kinds of DNA molecules are known to be more subject to mutation than others, resulting in faster mutation rates. This seems to be the case with the Y chromosome in human males.
Further criticism of the genetic argument for the replacement model has come from geneticists at Oxford University. They found that the human betaglobin gene is widely distributed in Asia but not in Africa. Since this gene is thought to have originated more than 200,000 years ago, it undercuts the claim that an African population of modern Homo sapiens replaced East Asian archaic humans less than 60,000 years ago.
Regional Continuity Model Arguments
Fossil evidence also is used to support the regional continuity model. Its advocates claim that there has been a continuity of some anatomical traits from archaic humans to modern humans in Europe and Asia. In other words, the Asian and European physical characteristics have antiquity in these regions going back over 100,000 years. They point to the fact that many Europeans have relatively heavy brow ridges and a high angle of their noses reminiscent of Neandertals. Similarly, it is claimed that some Chinese facial characteristics can be seen in an Asian archaic human fossil from Jinniushan dating to 200,000 years ago. Like Homo erectus, East Asians today commonly have shovel-shaped incisors while Africans and Europeans rarely do. This supports the contention of direct genetic links between Asian Homo erectus and modern Asians. Alan Thorne of the Australian National University believes that Australian aborigines ![]() share key skeletal and dental traits with pre-modern people who inhabited Indonesia at least 100,000 years ago. The implication is that there was no replacement by modern humans from Africa 60,000-40,000 years ago. However, the evidence does not rule out gene flow from African populations to Europe and Asia at that time and before. David Frayer, of the University of Kansas, believes that a number of European fossils from the last 50,000 years have characteristics that are the result of archaic and modern humans interbreeding.
share key skeletal and dental traits with pre-modern people who inhabited Indonesia at least 100,000 years ago. The implication is that there was no replacement by modern humans from Africa 60,000-40,000 years ago. However, the evidence does not rule out gene flow from African populations to Europe and Asia at that time and before. David Frayer, of the University of Kansas, believes that a number of European fossils from the last 50,000 years have characteristics that are the result of archaic and modern humans interbreeding.
Assimilation Model
It is apparent that both the complete replacement and the regional continuity models have difficulty accounting for all of the fossil and genetic data. What has emerged is a new hypothesis known as the assimilation (or partial replacement) model . It takes a middle ground and incorporates both of the old models. Gunter Brauer, of the University of Hamburg in Germany, proposes that the first modern humans did evolve in Africa, but when they migrated into other regions they did not simply replace existing human populations. Rather, they interbred to a limited degree with late archaic humans resulting in hybrid populations. In Europe, for instance, the first modern humans appear in the archaeological record rather suddenly around 45-40,000 years ago. The abruptness of the appearance of these Cro-Magnon people could be explained by their migrating into the region from Africa via an eastern Mediterranean coastal route. They apparently shared Europe with Neandertals for another 12,000 years or more. During this long time period, it is argued that interbreeding occurred and that the partially hybridized predominantly Cro-Magnon population ultimately became modern Europeans. In 2003, a discovery was made in a Romanian cave named Peştera cu Oase that supports this hypothesis. It was a partial skeleton of a 15-16 year old male Homo sapiens who lived about 30,000 years ago or a bit earlier. He had a mix of old and new anatomical features. The skull had characteristics of both modern and archaic humans. This could be explained as the result of interbreeding with Neandertals according to Erik Trinkaus of Washington University in St. Louis. Alan Templeton, also of Washington University, reported that a computer-based analysis of 10 different human DNA sequences indicates that there has been interbreeding between people living in Asia, Europe, and Africa for at least 600,000 years. This is consistent with the hypothesis that humans expanded again and again out of Africa and that these emigrants interbred with existing populations in Asia and Europe. It is also possible that migrations were not only in one direction--people could have migrated into Africa as well. If interbreeding occurred, it may have been a rare event. This is supported by the fact that most skeletons of Neandertals and Cro-Magnon people do not show hybrid characteristics.
It would be a mistake to assume that from the time modern Homo sapiens began migrating out of Africa that all people in that continent were modern humans. The video below presents evidence of pockets of archaic humans surviving in West Africa until at least 13,000 years ago.
Skull points to a more complex human evolution in Africa--Chris Stringer describes a problematical
human skull from Nigeria. This link takes you to a BBC website. Select the video at the top of the
article. To return here, you must click the "back" button on your browser program.
(length = 1 min, 45 secs)
Expansion Out of the Old World
Homo sapiens began migrating into the lower latitudes of East Asia by at least 70,000 years ago. Along the way, some of them interbred with archaic humans, including both Neandertals and Denisovans. Genetic markers from these archaic human populations are found in the gene pool of some Southern Chinese, New Guinean, and other Micronesian Island populations today. Homo sapiens from Southeast Asia travelled to Australia by 46,000 years ago and possibly as early as 60,000 years ago. Because Australia was not connected to Southeast Asia by land, it is probable that these first Australian Aborigines arrived by simple boats or rafts. Modern humans reached the Japanese Islands by 30,000 years ago or somewhat earlier. Around 35,000-30,000 years ago, Homo sapiens big game hunters moved into Northeastern Siberia. Some of them migrated into North America via the Bering Plain , or Beringia ![]() , by 20,000-15,000 years ago. Some Homo sapiens may have reached the Americas a bit earlier than this, but the evidence is still considered questionable by most paleoanthropologists. The Bering Plain intercontinental land connection appeared between Siberia and Alaska as a result of sea levels dropping up to 450 feet (137 m.) during the final major cold period of the last ice age. Until that time, all human evolution had occurred in the Old World.
, by 20,000-15,000 years ago. Some Homo sapiens may have reached the Americas a bit earlier than this, but the evidence is still considered questionable by most paleoanthropologists. The Bering Plain intercontinental land connection appeared between Siberia and Alaska as a result of sea levels dropping up to 450 feet (137 m.) during the final major cold period of the last ice age. Until that time, all human evolution had occurred in the Old World.
A consequence of human migrations into new regions of the world has been the extinction of many animal species indigenous to those areas. By 11,000 years ago, human hunters in the New World apparently had played a part in the extermination of 135 species of mammals, including 3/4 of the larger ones (mammoths, mastodons, giant sloths, etc.). Most of these extinctions apparently occurred within a few hundred years. It is likely that the rapidly changing climate at the end of the last ice age was a contributing factor. However, the addition of human hunters with spears to the existing top predators (mostly saber-toothed cats, lions, and dire-wolves) very likely disrupted the equilibrium between large herbivores and their predators. As a consequence there was a major ecosystem disruption resulting in the rapid decline of both non-human carnivores and their prey. Humans were very likely the trigger that set off this "trophic cascade". Unlike most other major predators, people survived by switching their food quest to smaller animals and plants.
F ollowing the arrival of aboriginal people in Australia and Polynesians in New Zealand there were similar dramatic animal extinctions. In both of these cases humans apparently were directly responsible for wiping out easily hunted species. Large vulnerable marsupials were the main victims in Australia. Within 5,000 years following the arrival of humans, approximately 90% of the marsupial species larger than a domesticated cat had become extinct there. In New Zealand, it was mostly large flightless birds that were driven to extinction by human hunters following their arrival in the 10th-13th centuries A.D.
It is sobering to realize that the rate of animal and plant extinction has once again accelerated dramatically. During the last century and a half, the explosion in our global human population and our rapid technological development has allowed us to move into and over-exploit most areas of our planet including the oceans. That exploitation has usually involved cutting down forests, changing the courses of rivers, pushing wild animals and plants out of farm and urban areas, polluting wetlands with pesticides and other man-made chemicals, and industrial-scale hunting of large land animals, whales, and fish. During the early 19th century, there were at least 40,000,000 bison roaming the Great Plains of North America. By the end of that century, there were only a few hundred remaining. They had been hunted to near extinction with guns. The same fate came to the African elephant and rhinoceros during the 20th century. Likewise, commercial fishermen have depleted one species of fish after another during the last half century. Governments have had to step in to try to stem the tide of these human population effects on other species. However, they have been only marginally successful. The World Conservation Union conservatively estimates that 7,266 animal species and 8,323 plant and lichen species are now at risk of extinction primarily due to human caused habitat degradation. The endangered list includes 1/3 of all amphibian species, nearly 1/2 of the turtles and tortoises, 1/4 of the mammals, 1/5 of the sharks and rays, and 1/8 of the birds. This list does not include the many millions of species that are still unknown to science. It is likely that most of them will become extinct before they can be described and studied.
People Today
Are we genetically different from our Homo sapiens ancestors who lived 10-20,000 years ago? The answer is almost certainly yes. In fact, it is very likely that the rate of evolution for our species has continuously accelerated since the end of the last ice age, roughly 10,000 years ago. This is mostly due to the fact that our human population has explosively grown and moved into new kinds of environments, including cities, where we have been subject to new natural selection pressures. For instance, our larger and denser populations have made it far easier for contagious diseases, such as tuberculosis, small pox, the plague, and influenza to rapidly spread through communities and wreak havoc. This has exerted strong selection for individuals who were fortunate to have immune systems that allowed them to survive. There also has been a marked change in diet for most people since the end of the last ice age. It is now less varied and predominantly vegetarian around the globe with a heavy dependence on foods made from cereal grains. It is likely that the human species has been able to adapt to these and other new environmental pressures because it has acquired a steadily greater genetic diversity. A larger population naturally has more mutations adding variation to its gene pool simply because there are more people. This happens even if the mutation rate per person remains the same. However, the mutation rate may have actually increased because we have been exposed to new kinds of man-made environmental pollution that can cause additional mutations.
It is not clear what all of the consequences of the environmental and behavioral changes for humans have been. However, it does appear that the average human body size has become somewhat shorter over the last 10,000 years, and we have acquired widespread immunity to the more severe effects of some diseases such as measles and influenza.
Finally, c an we say what direction human evolution will take in the future? This is a fascinating question to consider but impossible to answer because of innumerable unknown factors. Though, it is certain that we will continue to evolve until we reach the point of extinction.
NEWS: Maanasa Raghavan et al. reported on November 20, 2013 ( http://www.nytimes.com/2013/11/21/science/two-surprises-in-dna-of-boy-found-buried-in-siberia.html?pagewanted=1&_r=1 and http://www.nature.com/nature/journal/vaop/ncurrent/full/nature12736.html ) that a 24,000 year old child skeleton from Mal�ta near Lake Baikal in eastern Siberiashared major DNA markers with Western Europeans and 1/4 of living Native American. A 17,000 year old adult skeleton from Siberia also had genetic markers indicating European origin. This indicates that ice age Homo sapiens from Europe migrated much farther beyond that continent than commonly believed. It also implies that the New World native populations owe their ancestry not only to East Asians but to Europeans as well .
Copyright � 1999-2013 by Dennis O'Neil. All rights reserved.
illustration credits
Source: https://www2.palomar.edu/anthro/homo2/mod_homo_4.htm
0 Response to "What Does the Multiregional Continuity Thesis Propose"
Post a Comment